JMS Pearce
London, United Kingdom
Early evolutionary theorists noted that the evolution of the brain, its structural organization, and microscopic structure appeared to develop concurrently with human anatomy through the process of evolution. This understanding of the evolution of the brain was heavily influenced by both the theories of phylogeny, which discusses this evolutionary lineage of a species, and ontogeny, which purports that the development or progression of a species is reflected or recapitulated during the embryonic development of the individual.
The presumption that the evolution of the central nervous system mirrors that of anatomical evolution at first seemed quite logical. For instance, the most complex brains include the arthropods (insects and crustaceans), cephalopods (octopuses, squids, and molluscs), and vertebrates. Arthropod and cephalopod brains develop from twin nerve cords. Vertebrate brains develop from a single dorsal nerve cord; and complex vertebrates, such as mammals, have a cerebrum with a six-layered neocortex (neopallium) and many convolutions that provide a larger area for a greater number of neurons within a compact volume. Primitive vertebrates—fish, reptiles, and amphibians have fewer than six layers of neurons in their cerebral cortices. However, more recent developments in the understanding of human evolution tend to refute this rather simplistic understanding of the human brain.
While Aristotle had earlier noted the anatomical variability marked in humans “to whose share had fallen not mere life, but life of high degree,” understanding of the brain’s evolving morphology was advanced by microscopy. Ramon y Cajal first elaborated the comparative histology of animal neuronal systems, which, in describing the variation within the species, questioned the link between ontogeny and phylogeny and the evolution of the nervous system. Darwin, in turn, was occupied by the enormous diversity of form within species, from which he deduced the Theory of Natural Selection (Table 1: Timeline). Work by these theorists and others have laid the framework for the lasting controversy surrounding the evolutionary theories of ontogeny and phylogeny and made way for the discussion of Neural Darwinism as the means by which the human nervous system has evolved to its present form.
Early evolutionary discoveries
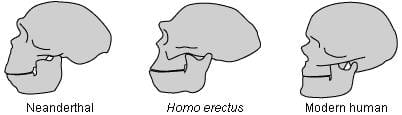
Humans share a common ancestor with African apes, whose fossils were found in Africa, Asia and Europe. Since primate and human skulls are similar in terms of brain size, shape and development, their evolutionary forms (Fig. 1) have been studied in order to shed light on human evolution; indeed the discovery of the Neanderthal skull marked the beginning of paleoanthropology. It was discovered in Gibraltar in 1848, three years before Darwin’s Origin of Species.1 Apart from Homo neanderthalis, not until the last hundred years were fossils discovered.
In 1925, Raymond Dart, one of the pioneers of paleoanthropology, described Australopithecus africanus. The australopithecines, the hominid family to which A. africanus belongs, first appeared roughly 4.2 million years ago; they are the immediate ancestors of the genus Homo sapiens sapiens. Dart’s Taung Child, the only discovery of this type that he made, was found in South Africa. It had a tiny skull and brain (410 ml.); the skull was rounded, with short ungrooved canine teeth, the brain more like a human than a chimpanzee or gorilla brain. Dart thought the Taung Child was a bipedal human ancestor, intermediate between apes and humans. Mary Leakey later discovered Paranthropus boisei in 1959 in Tanzania. At first classed with the Australopithecus genus, this hominid was later renamed P. boisei, and probably evolved 2.3 to 1 million years ago along a path unrelated to early humans. Each of these discoveries contributed to the development and understanding of the evolutionary theory as it stands today.
Ontogeny recapitulates phylogeny
In 1866, following the discoveries of the Neanderthal skull, Ernest Haeckel (1834–1919), a brilliant German marine biologist, introduced the word phylogeny, the line of descent of all organic beings.2 Phylogeny can be vertical, showing changing morphology from earliest to latest species, or horizontal, where different living organisms develop homologous traits at the same time. Early theorists readily applied the concept of phylogeny to the brain since both ontogeny and phylogeny reveal progression with evolutionary time. For instance the evolutionary progression of mammals from their aquatic ancestors reveals increasing encephalization from amphibians to reptiles and so on.
Haeckel’s scheme took phylogeny a step further, theorizing that animals advanced to new levels by adding stages to existing embryonic development, leading to the then widely popular recapitulation theory of evolution. Ontogeny was the idea that the development or progression of a species is reflected or recapitulated during the embryonic development of the individual. For example, the human embryo with gill slits in the neck was believed by Haeckel to signify not only a fishlike ancestor, but to represent a fishlike stage in human embryogenesis. Hence emerged Haeckel’s Biogenetic Law that embryonic stages of development move through phases of phyletic evolution; in other words, “ontogeny recapitulates phylogeny.” Ancestral adult stages were “accelerated” into the ontogeny of the descendants. The idea was seductive and widely influential, reproduced in many 19th and 20th century European texts:
The earliest characters of the embryonic head or of its equivalent in all Vertebrates, point to its being phylogenetically, the most ancient part of the body, and serve as a finger-post to the path of development of the Vertebrate body.3
Even as late as the 1960s, Benjamin Spock’s best-selling book on childcare revisits recapitulation theory:
Each child, as he develops, is retracing the whole history of mankind, physically and spiritually, step by step. A baby starts off in the womb as a single tiny cell, just the way the first living thing appeared in the ocean. Weeks later, as he lies in the amniotic fluid in the womb, he has gills like a fish.4
Not all scientists subscribed to Haeckel’s theory, however, and some scientists had long rejected the concept. Although Haeckel was an enthusiastic supporter and correspondent of Darwin, Darwin did not particularly support the concept of ontogeny, but stated:
Nothing can be effected, unless favourable variations occur … I do believe that natural selection will always act very slowly, often only at long intervals of time, and generally on only a very few of the inhabitants of the same region at the same time. I further believe, that this very slow, intermittent action of natural selection accords perfectly well with what geology tells us of the rate and manner at which the inhabitants of this world have changed … I am convinced that natural selection has been the main but not the exclusive means of modification.5
The famous Estonian, Karl Ernst Von Baer (1792–1876), physician, zoologist and anthropologist, had taken an opposite view in 1828, 40 years before Haeckel:
Every embryo of a given animal form, instead of passing through the other forms, rather becomes separated from them. . . . Fundamentally, therefore, the embryo of a higher form never resembles any other form, but only its embryo.6
Von Baer had discovered the mammalian egg and the formation of the germ layers whence embryonic organs develop. While this discovery laid the basis for both ontogeny and embryology, opposition to and, ultimately, rejection of the notion of recapitulation was to follow.
Laying the foundation for the Neuron Doctrine
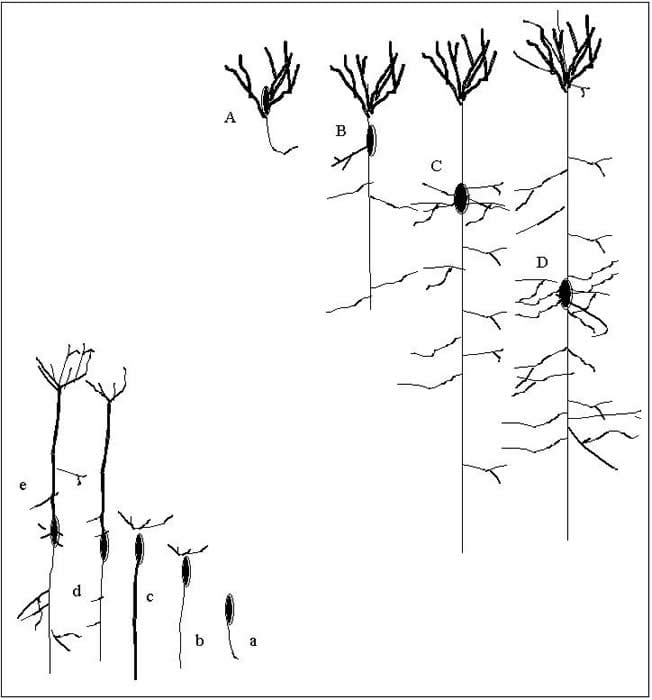
Above: A. frog, B. lizard, C. rat, D. man. Below: a. neuroblast without protoplasmic stalk, b. beginning stalk of terminal plume, c. further developed, d. appearance of axonal collaterals, e. protoplasmic processes and stalk.
Santiago Ramon y Cajal (1852–1934), a Spanish histologist and Nobel laureate, was cautious to avoid controversy; nonetheless his work provides fascinating histological detail of the changing morphology of the brain’s cortical pyramidal neurons. His monumental work7 described the cytological foundations that allowed for the developments of the neuron doctrine, a now well-accepted idea that the nervous system is comprised of individual cells. From portrayals of the complex primate configurations (Fig. 2) to primitive fish neurons, he observed:
[As] one descends the scale … the dendritic apparatus appears to be less differentiated, and the collaterals of the axons less numerous, less long, and less branched …. as the cerebral cortex increases the dendrites and the collaterals of the axons of the pyramids become proportionately longer and more branched.8
Cajal did not suggest a new principle, but his detailed descriptions permitted the systematic comparison of neurological structure between species and humans. He went on to propose a correlation between microscopic neuronal structure and intellect, later supported by fossils that shed light on the skull, brain size and convolutional patterns of human ancestors. How this process evolved Darwin eventually related to natural selection, laying the way for the theory of Neural Darwinism.
Recapitulation refuted
The 19th and 20th century ideas with which recapitulation theory evolved suggested somewhat teleologically that tissues, organs and individual animals gradually modified their forms through the process of natural selection over millions of years in order to adapt to changing environments, food sources and predators. This theory, however, was opposed not only by the Church, but also by scientists, including Von Baer, due to unexplained gaps in the evolutionary process. Even Darwin admitted that a gradual evolutionary process did not correspond well to the rarity of intermediate forms, or missing links.
In the middle of the 20th century, Stephen Jay Gould partly resolved this issue through the development of the punctualistic theory, which speculates that species have long periods of stasis and short periods of divergence. Punctuated equilibria, as the theory is otherwise known, proposed sudden stepwise9 evolutionary changes in the morphology of developing new species, rather than the gradual changes in the proposed original theory of natural selection,10 thereby rejecting the “ontogeny recapitulates phylogeny” theory:
Sudden appearance and stasis shall dominate the fossil record. All major theories of speciation maintain that splitting takes place rapidly in very small populations. … A new species can arise when a small segment of the ancestral population is isolated at the periphery of the ancestral range. Large, stable central populations exert a strong homogenizing influence. New and favorable mutations are diluted by the sheer bulk of the population through which they must spread. … Thus, phyletic transformation in large populations should be very rare—as the fossil record proclaims. But small, peripherally isolated groups … live as tiny populations in geographic corners of the ancestral range. Selective pressures are usually intense because peripheries mark the edge of ecological tolerance for ancestral forms. Favorable variations spread quickly. Small peripheral isolates are a laboratory of evolutionary change.
Gould’s theory is now widely if not universally accepted. With the refutation of the recapitulation theory, new theories have also emerged to explain the development of the central nervous system.
Experimental embryology and Neural Darwinism
Theories of recapitulation eventually gave way with the emergence of embryology, which attempts to unlock the genetic code for human morphological change through the study of how genes regulate the development of the embryo. Mendel’s work provided these scientists with a method for applying the theory of natural selection to the propagation of certain genetic characteristics. The study of embryonic brain development has revealed that its anatomical connections, called primary neuron groups, are created not only by genetic factors (which regulate such things as cell shape), but also by epigenetic events (heritable changes that occur through the activation or inhibition of genes, without change in the sequence of nuclear DNA).

Epigenetic events regulate cell division, movement, differentiation and neuronal death. The diversity of nerve cell connections regulated through these epigenetic events results in an innumerable variety of neuronal groupings, which are unique to every animal. Axons and dendritic connections are therefore similar in animals of the same species, but are subject to individual variation. It is this variation that permits adaptation to evolutionary change. The formation of neuronal groups has been linked with adaptive behavior such as the acquisition of an upright posture and bipedalism.
In an extrapolation from Cajal’s comparative neuronal morphology, Edelman, winner of the 1972 Nobel Prize, asserted that this variation in structure is caused by selective mechano–chemical events—a mechanism of natural selection. Cell and substrate adhesion molecules regulate these epigenetic events, which in turn determine cell division, movement, differentiation and cell death. Cell adhesion molecules guide the development of an animal’s overall shape and form as well as the construction of the nervous system. Genetic mutation may also alter how the expression of these molecules is regulated, leading to evolution within specific brain areas. For instance, the precursor gene for the neural cell adhesion molecule gave rise to the entire molecular system of adaptive immunity. Apart from mutations, the elements of the brain appear to organize and adjust their connections in response to environmental demands.
The partitioning of the early vertebrate hindbrain also seems to contribute to the evolution of neural networks, which in turn develop as independent modules, called somites or metameres, which evolve independently.12 However, hindbrain segmentation occurs exclusively in the embryo; neurons and synapses are reconfigured and reintegrated in fetal and postnatal stages. The brain continues to develop a huge diversity of circuits during fetal growth and throughout life. According to the theory of Neural Darwinism, the nervous system develops synaptic arrangements, or connections, from one nerve cell to another, which are strengthened or weakened depending on environmental stimuli, resulting in a huge number of “repertoires of variance.” Thus all brains, even those of identical twins, are unique.
Edelman’s re-entry theory, the final tenet of Neural Darwinism, posits the origin of consciousness as deriving from projections of sensory or motor information from the thalamocortical system through the electrical stimulation of massive pathways of parallel, dynamic reciprocal neural connections. It is regular electrical stimulation of these neural connections that links diverse neural networks, or maps, within the brain to produce action or function.13 The natural selection of the various neural circuits, occurring through the interaction of environmental exposure and anatomy in the process of re-entry, results in the wealth and diversity of conscious experience.
Conservation of genetic mechanisms: A common evolutionary origin?
The organization of the human cortex has evolved over 4 million years.14 The neocortex is a thin, extended, convoluted sheet with a surface area of about 2600 cm2, and thickness of 3–4 mm. It contains up to 28 x 109 neurons and approximately the same number of glial cells. Cortical neurons are connected by a vast number, about 1012, of synapses. The cortex is organized horizontally and vertically into groups of cells linked by synapses. Despite current theories of evolution and scientific discovery, we still understand neither its intrinsic operations, nor how these elementary units in different cortical areas are linked dynamically in the systems of the neocortex. The many and varied differences in morphology and the diversity of nerve cell circuits in fetuses are present both in invertebrates and vertebrates. Whether they indicate a common ancestry or a divergence in genera at intervals in the long evolutionary timescale is still uncertain.
Observing the expression of genes between widely differing genera leads to contradictory conclusions about the origin of each species. For instance, there are obvious anatomical differences between the gastroneuralia (arthropods, annelids, molluscs), which have a ventral nerve cord containing somite ganglia, and the notoneuralia (all chordates), which have a dorsal nerve cord derived from dorsal neuroectoderm. Such differences in embryonic topography of the nervous system15suggest a differing, independent evolutionary origin. However, molecular genetic studies show remarkable similarities in the expression of regulatory genes that control neural embryogenesis in both insects and mammals.16 This strongly suggests that these genetic mechanisms are conserved in brain embryogenesis for biological advantages for the organism and a shared evolutionary origin.17
The last common ancestor to the diversity of modern bilaterian species (animal with symmetry of body parts), seen in the early Precambrian (period >4500 million years ago), had similar characteristics to modern bilaterians.18 This supports the conservation of genetic mechanisms that have been discovered across the bilateria. For instance, because the genetic expression leading to differentiation and growth of tissues during the development of various organs and body parts is similar in both insects and vertebrates, it suggests that the origin of these genetic traits is shared. Thus, the eye, heart, gut, peripheral and central nervous systems seem to share similar genetic origins across species.
Conclusion
The history of evolutionary theory provides as many unanswered questions as discoveries, with each new theory providing a new and necessary understanding, while dismissing popular theories of the past. While questions still remain regarding the origin of genetic traits shared across diverse species, Haeckel’s initially attractive “ontogeny recapitulates phylogeny” theory has largely been disproved, despite its persistence in popular dialogue. Discoveries regarding the changing complexity of the brain, so well illustrated by Cajal, may have laid the foundations for the Neural Doctrine and Neural Darwinism, but, at the same time, they are not necessarily indicative of progress. As Darwin stated:
[A]daptation to changing local environments, which are randomly moving through time. … But although evolution means progress, there is no principle of general advance in that.19
Populations adapt to their current surroundings, but do not necessarily become better in any absolute sense. A change that is successful for survival at one time may be unsuccessful at another. In addition, despite what appears to be increasing complexity in terms of neurological development, organisms as a rule become decreasingly complex in terms of form as they evolve. Early Homo neanderthalis, for instance, had a 1200–1700 cm3 skull capacity—greater than modern humans. H. neanderthalis, which shared a common ancestor with modern humans, also had an occipital bun, mastoid crest, prominent supraorbital ridge but almost no chin—“primitive” features that disappeared in Homo sapiens. Whether complexity per se is a sign of advance or superiority remains uncertain, making these conventional terms seem arbitrary and abstract.
Despite our current incomplete understanding of human evolution and the corresponding development of the nervous system, one is left only to marvel at the evolutionary path that has led to human consciousness and its seemingly endless diversity; for as Gould says in the Evolution of Life on Earth:
Humans arose, as a fortuitous and contingent outcome of thousands of linked events, any one of which could have occurred differently and sent history on an alternative pathway that would not have led to consciousness.20
Table 1. Timeline
|
1848
|
Early skulls and brain: beginning of Paleoanthropology
|
|
1859
| Darwin’s On the Origin of Species by Means of Natural Selection. Natural selection has been the main but not the exclusive means of modification. |
|
1866
| Haeckel’s “Ontogeny recapitulates Phylogeny” theory is developed: animals advance to new levels by adding stages to existing embryonic development. |
|
1866–1873
| Mendel’s genetic mechanism for natural selection is formulated. |
|
1880–1910
| Cajal’s comparative anatomy of neurones is developed, noting that as the cerebral cortex increases, the dendrites and the collaterals of the axons become proportionately longer and more branched. |
|
1948
| Edelman’s Neural Darwinism: variation in structure is caused by selective mechano–chemical events regulated by “cell and substrate adhesion molecules” that determine cell division, differentiation, and cell death. |
|
1977
| Gould and Eldredge publish their theory punctuated equilibria, proposing sudden stepwise evolutionary changes in the morphology of developing new species |
|
1990s
| Conservation of genetic mechanisms: a common evolutionary origin? |
Notes
- Darwin, C. (1859). On the origin of species by means of natural selection, or the preservation of favoured races in the struggle for life. London: John Murray.
- Haeckel, E. (1866). Generelle morphologie der organismen (Vols.1-2). Berlin: G. Reimer.
- Gegenbaur, C. (1878). Elements of comparative anatomy. (F.J. Bell & E.R. Lankester, Trans.). London: Macmillan.
- Spock, B. (1968). The common sense book of baby and child care (2nd ed.). New York: Meredith Press.
- Darwin, C. (1859). On the origin of species by means of natural selection, or the preservation of favoured races in the struggle for life. London: John Murray.
- Von Baer, K.E. (1853). Fragments relating to philosophical zoology. (T. H. Huxley, Trans.). In A. Henfrey and T. H. Huxley (Eds.), Scientific memoirs, selected from the transactions of foreign academies of science and from foreign journals (pp. 176-238). London: Taylor and Francis. (Original work published in 1828).
- Cajal, S. R. (1899, 1904). Textura del sistema nervioso del hombre y de los vertebrados (Vols. 1-2).. Madrid: Imprenta y Librería de Nicolás Moya.
- Cajal, S. R. (1892). El nuevo concepto de la histología de los centros nerviosos. Conferencias dadas en la Academia y Laboratorio de Ciencias Médicas de Cataluña. Revista de ciencias medicas, 18, 457-476.
- Gould S. J., & Eldredge, N. (1977). Punctuated equilibria: the tempo and mode of evolution reconsidered. Paleobiology, 3, 115-151.
- Gould S. J. (1977). Ontogeny and phylogeny. Cambridge, Mass: Belknap Press.
- Gould S. J. (1980). The episodic nature of evolutionary change. The panda’s thumb: More reflections in natural history (pp. 182-184). New York: W. W. Norton.
- Edelman, G. M. (1993). Neural Darwinism: Selection and reentrant signalling in higher brain function. Neuron, 10, 115-125.
- Lumsden A. (1990). The cellular basis of segmentation in the developing hindbrain. Trends in Neurosciences, 13, 329-35.
- Mountcastle, V. B. (2003). Introduction. Cerebral Cortex, 13, 2-4.
- Arendt, D. & Nübler-Jung, K. (1999). Comparison of early nerve cord development in insects and vertebrates. Development, 126, 2309-2325.
- Reichert, H. Conserved genetic mechanisms for embryonic brain patterning. (2002). International Journal of Biology, 46, 81-7.
- Thor, S. (1995). The genetics of brain development: Conserved programs in flies and mice. Neuron, 15, 975-977.
- Erwin, D. H. & Davidson, E. H. (2002). The last common bilaterian ancestor. Development, 129, 3021-3032.
- Koch, J. (1995, December 31). [Interview with Stephen Jay Gould]. Boston Globe Magazine.
- Gould, S. J. (1995). The pattern of life’s history. In J. Brockman (Ed.), The third culture: Beyond the scientific revolution (pp. 51-73). New York: Simon & Schuster.
- Gould S. J. (1994, October). The evolution of life on the Earth. Scientific American, 85-86.
, MD, FRCP (London) is Emeritus Consultant Neurologist in the Department of Neurology at the Hull Royal Infirmary, England.
Highlighted in Frontispiece Volume 18, Issue 2 – Spring 2026 and Volume 2, Issue 2 – Spring 2010