James L. Franklin
Chicago, Illinois, United States
This is part two of a two-part article. See part one.
Color Blindness
With this background on color vision, we return to the subject of color blindness. A familial component, what we now recognize as inherited or genetic, was evident in the earliest reports of color blindness. Captain Joseph Huddart’s 1777 letter to Rev. Joseph Priestley mentions that the shoemaker Thomas Harris had “two brothers in the same circumstance as to sight, and two other brothers and sisters who, as well as their parents had nothing of this defect.” In his paper of 1794, John Dalton also noted familial occurrences of defective color vision including that of his own brother.
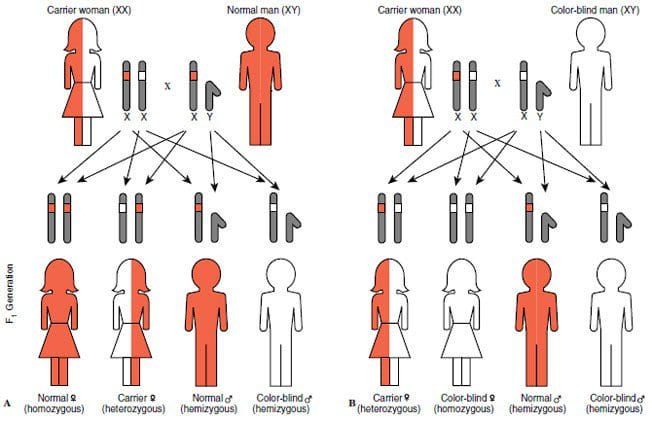
Color blindness is the first human genetic trait to be linked to a specific chromosome. The Swiss ophthalmologist Johann Friedrich Horner (1831 – 1886), known to generations of medical students for “Horner’s Syndrome” and its association with tertiary syphilis, presented the first scientific account of the hereditary transmission of Daltonism. He recognized that color-blind fathers have color-normal daughters who are in turn mothers of colorblind sons.[i] Mothers who carry the gene on one of their X chromosomes have normal vision.
Women also may inherit color defective vision but with a far lower frequency. One of every two daughters resulting from the union of a colorblind father and a mother who is a carrier of the colorblind gene will have defective color vision. [Figure 4]
Since the methods for testing defective color vision during the nineteenth century were not standardized and only partially understood, much of the data gathered on individuals with color defective vision was confusing. In 1895, Johannes Adolf von Kries (1853 – 1928), a German physiologist and psychologist and disciple of Herman Helmholtz, coined the terms used today in classifying defective color vision. Based on the trichromatic theory of color vision, he classified subjects as having protanopia, deuternopia and tritanopia. These terms refer to the missing cone receptor; protanopia, first (red), deuteranopia, second (green) and tritanopia, third (blue). [Table 1]
Table 1: Prevalence of Color Blindness
Tritanomaly (blue deficient: S cone defect)0.0001%0.0001%
| Types of Color Blindness | Males | Females |
|---|---|---|
| Dichromacy | 2.4% | 0.03% |
| Protanopia (red deficient: L cone absent) | 1.3% | 0.02% |
| Deuteranopia (green deficient: M cone absent) | 1.2% | 0.02% |
| Tritanopia (blue deficient: S cone absent) | 0.001% | 0.03% |
| Anomalous Trichromacy | 6.3% | 0.37% |
| Protanomaly (red deficient: L cone defect) | 1.3% | 0.02% |
| Deuteranomaly (green deficient: M cone defect) | 5.0% | 0.35% |
The photo pigments in each of the three cone receptors are genetically determined, chemically distinct molecules that differ from each other by only a few amino acids. These small differences account for their variable sensitivity to light across the visible spectrum.
Humans with normal color vision recognize color by processing signals from the three types of cone receptors and are referred to as ‘trichromats,’ or as having ‘trichromatic vision.’ Individuals totally lacking one of the primary visual pigments have ‘dichromatic’ vision. Dichromats match colors by processing the differences between two color receptors. Classification is further complicated because many individuals with defective color vision have inherited genes that alter the sensitivity of one of the three cone photoreceptors, most frequently red or green. In these instances, the receptor is still functional and the affected individual still has trichromatic vision. They are classified as having ‘Anomalous Trichromacy’ and sub-classified as protanomaly, deuteranomaly and tritanomaly. Individuals with ‘Anomalous Trichromacy’ experience a richer palette of colors than dichromats and are less likely to realize they have abnormal color vision.
Evolutionary Trait
The question may be asked why defective color vision, a sex- linked recessive trait, persists in the human genome with a frequency that rises to eight percent of the male Caucasian population? In the classic example of sickle cell anemia, also inherited as a recessive trait, carriers of the sickle cell gene are more resistant to infection by the malarial parasite than non-carriers. This confers a survival advantage for carriers of the sickle cell gene and explains the persistence of an otherwise deleterious gene. A Darwinian is led to ask, does defective color vision or its carrier state confer a natural selective advantage or is it survival neutral?
The answer seems to require a broader view of the evolution of color vision. The photoreceptor pigments in the rods and cones have ancient origins within the animal kingdom. The gene for the blue cones is the most ancient and pre-dates the entire mammalian line.2 Primates of South America, New World monkeys, broke away from African, Old World monkeys about sixty-three million years ago. The New World monkeys uniformly have dichromatic vision while Old World monkeys, with whom we share a common ancestor, evolved trichromatic vision. Our ancestors managed to duplicate the X-linked receptor gene and acquire separate classes of red and green cones. There are lines of evidence to suggest that primate trichromatic vision is as young as 30 – 40 million years old. As one author has noted, “In evolutionary terms, that is a last-minute job. No wonder it goes wrong so often.”3
None of the Old World primates studied thus far has been found to have a defect comparable to human color blindness, that is dichromatic vision. This suggests in an arboreal habitat, red-green color deficiency would be a negative survival factor, such as difficulty recognizing ripe fruit in the canopy of the forest. Once our ancestors left the forests on their long march to civilization, survival pressure against the mutation eased.
Could color blindness or its carrier state confer a survival or sexual selective advantage? There are poorly documented references suggesting that during WW II, color-blind airmen made better bombardiers due to their ability to see through enemy camouflage— hardly a factor of evolutionary significance. Nor has anyone mustered evidence that abnormal color vision might be sexy. John Dalton and Quakers in general were known to have rather drab taste in dress.
What about the demographics of color blindness? Dr. Shinobu Ishihara, the Japanese Ophthalmologist, who created the Ishihara color test, was born in Tokyo in 1879. He was trained in ophthalmology in both Tokyo and Germany. With the outbreak of WW I, he was asked by the Japanese military to develop a color vision test to screen conscripts for defective color vision. It is remarkable that he hand painted his first set of test plates. The test consists of a circular field in which dots of various sizes and hues are evenly distributed. [Figure 5] Within the field shapes (numbers) were created that could only be identified on the basis of color. The principle on which the test is based is called pseudo-isochromaticism where two or more colors appear equivalent but in fact are distinct.

Introduction of the Ishihara plates made it possible to perform larger scale population surveys for defective color vision. Across the globe, zones of high, intermediate and low prevalence have been mapped. Europe and North America show a high prevalence approaching eight percent while India and China belong to the intermediate regions of four to six percent. Eskimos in the arctic as well as tribal peoples in the equatorial forests of South America, Africa and New Guinea have a low prevalence of less than four percent. In South America, strong differences exist between isolated rural areas and urban industrial regions with a gradient of color blindness from 1.6% in semi-nomadic Andean shepherds to eight percent in industrial port cities in Chile. A survey of the frequency of color blindness among 60,000 R.A.F. men in 1943 pointed to a higher percentage in men from the western areas of Britain corresponding to regions invaded by Angles, Saxons and Jutes during the fifth and sixth centuries.4 The distribution of defective color vision across the globe has been interpreted in a number of different ways, but one is left with the impression that color blindness might be a serious handicap in primitive cultures. With the agricultural revolution, selective pressure relaxed allowing mutant genes to persist.
John Dalton’s Eyes – Part Two
We now return to John Dalton’s eyes and those desiccated fragments that have been held by the Manchester Literary and Philosophical Society. In 1995, one hundred and fifty years after his death, British scientists published a remarkable study in molecular biography, “The Chemistry of John Dalton’s Color Blindness.” Analysis of the DNA in samples taken from the fragments of Dalton’s eyes revealed that Dalton had a single LW (Long Wave – red) gene and lacked the MW (Middle Wave – green) gene. Dalton had dichromatic vision and was a deuteranope. Their findings contradicted the traditional belief that Dalton was a protanope with a foreshortening of the red end of the spectrum. Thomas Young was one source of the belief that Dalton lacked “those fibers of the retina that are calculated to perceive red.” He was influenced by Dalton’s description of the solar spectrum: “that part of the image which others call red appears to me little more than a shade or defect of light.”
In 1968, W.D. Wright, a professor of applied optics at the Imperial College of Science and Technology, reviewed Dalton’s self-observations on color concluding that they unequivocally support the fact that he was a protanope. The authors of “The Chemistry of John Dalton’s color Blindness (1995),” explain that to a deuteranope, the “red part of the spectrum will look dim because the regions that look yellow, orange and red to normal observers are for him all of the same hue, but red is of lower luminosity than the yellow and orange regions to which it is juxtaposed.” They further point out that Sir John Hershel and Sir David Brewster who questioned Dalton found that he did not see the red end of the spectrum as foreshortened. Their analysis of Dalton’s self-observations finds them to be consistent their DNA evidence that he was a deuteranope.
On Monday, August 12, 1844, the municipality of Manchester made amends for their tardy recognition of his attainments by giving Dalton a public funeral. The body “lay instate” for three days in the Town Hall and was visited by over forty thousand people. A funeral cortege of approximately one hundred carriages and four hundred police were on duty along the route to Ardwick Cemetery. All public bondies in Manchester were represented. Dalton’s grave is a large vault covered with heavy flagstones. The tomb is of polished granite with the name “John Dalton” and the dates of his birth and death.5 The cemetery in 1944 was obscured by buildings and at the time known to few Mancurians. In the 1960s the site was converted to a sports fields. The granite slab with Dalton’s name lies next to a statue of John Dalton located outside the John Dalton Building of the Manchester metropolitan Museum.
John Dalton, requiescat in pace!
Notes
- “Horner’s Syndrome” (Ptosis – unilateral drooping of an eyelid, meiosis – a small pupil, enophthalmos – a sunken or depressed eyeball and unilateral anhydrosis – absence of facial sweating on the affected side).
- The genes encoding ‘blue’ cones are located on chromosome 7, an autosome and have a different evolutionary history than the red-green cones. This means tritanopia and tritanomaly are not sex-linked traits and women and men are equally affected. They exist with an approximate prevalence of one in 10,000 individuals.
- Simon Ings, “A Natural History of Seeing,” W.W. Norton & Company, 2007, p. 259.
- Vernon, P.E. and Stracker, R.A. Distribution of colour blindness in Great Britain, Nature, 152:690,1960
- T. Lenton Elliott, John Dalton’s Grave, The Journal of Chemical Education, 1953:30 (11);569
Selected Bibliography on Color Vision and Color Blindness (Books):
- Cruz-Coke, Ricardo (1970) Color Blindness: An Evolutionary Approach, Charles C Thomas, USA
- Gregory, R.L. (1978) Eye and Brain: the psychology of seeing, Third Edition, MGraw Hill, NY Toronto
- Ings, Simon (2007) A Natural History of Seeing: The art & Science of Vision, W.W. Norton & Company, New York
- Valberg, Arne (2005) Light Vision Color, John Wiley & Sons, England
- Wasserman, Gerald S. (1978) Color Vision: An Historical Introduction, John Wiley & Sons, Inc.
- Wright, W.D. The Rays are not Coloured: Essays on the science of vision and colour, (1968), Chapter 5 The Unsolved Problem of Daltonism, pp. 67-87, American Elsevier Publishing Company, New York.
Articles of Historical Interest from Scientific American:
- Jeremy Nathans, The Genes of Color Vision, Scientific American: 42-49, February 1989
- Edward MacNichol, Jr. Three-Pigment Color Vision. Scientific American 211:48-56, December 1964
- Richard H. Masland, The Functional Architecture of the Retina,
- W.A.H. Rushton, Visual Pigments and Color Blindness. Scientific American 232: 64- 74, March 1975
- W.A.H. Rushton, Visual Pigments in Man, Scientific American, 207: 120-132, November 1962
- George Wald, Life and Light, Scientific American 201:92-108, October 1959
Biography:
- Michel Meulders (translated and edited by Laurence Garey) , Helmhotz: From Enlightenment to Neuroscience, The MIT Press, Cambridge Massachusetts, 2010
- Basil Mahon, The man who Changed Everything: The Life of James Clerk Maxwell, John Wiley & Sons, Ltd. England, 2004
- Andrew Robinson, The Last Man who Knew Everything: Thomas Young, The Anonymous Genius Who Proved Newton Wrong and Deciphered the Rosetta Stone, Among Other Surprising Feats, A Plume Book, USA, 2007
Articles on Color Blindness:
- Hunt, DM et. al. The Chemistry of John Dalton’s color blindness. Science 1995: 232: 203-210
- Fishman, Gerald A. John Dalton: Though in Error, He Still Influenced Our Understanding of Congenital Color Deficiency. Opthalmic Genetics2008:29:162-165
- Snyder, Charles, The Eyes of John Dalton, Archives of Ophthalmology, 1962; 67:671- 673 1 Portions of this paper were read at a meeting of The Chicago Literary Club on March 21, 2016
JAMES L. FRANKLIN is a gastroenterologist and associate professor emeritus at Rush University Medical Center. He also serves on the editorial board of Hektoen International and as the president of Hektoen’s Society of Medical History & Humanities.